多杀菌素类杀虫剂研发进展
| 2025-12-01 | 来源:农药资讯网 | 作者: | 【字体:大 中 小】 |  |
原标题:多杀菌素类杀虫剂及其作用靶标的研究综述
来源:《中国植保导刊》2025年第8期
作者:孙丽娜1 赵毅渊1 张怀江1 闫文涛1 岳强1 窦术英1 孟威2 李维根2 仇贵生1(1.中国农业科学院果树研究所;2.辽宁省植保植检总站)
生物农药主要包括微生物农药、生物化学农药、植物源农药、抗生素农药,多杀菌素类农药属于微生物农药,有效成分由土壤放线菌刺糖多胞菌(Saccharopolyspora spinosa)发酵产生,具有复杂的化学结构和广泛的杀虫谱。作为一类高效、安全的生物农药,多杀菌素类杀虫剂在农林害虫、储粮害虫、卫生害虫及牲畜寄生虫的防治中发挥着重要作用。
01 多杀菌素类杀虫剂结构特点与作用方式
1.1 结构特点
1929年青霉素的发现开启了天然产物研究的黄金时代,目前已从微生物中发现超过6,000种具有天然活性的产物,包括常用杀虫剂阿维菌素和多杀霉素等。20世纪80年代,美国礼来公司从维尔京群岛一个废弃甘蔗加工厂附近的土壤中分离得到刺糖多胞菌,其发酵液的主要杀虫活性成分为多杀菌素A和多杀菌素D。后续开发出活性更高的乙基多杀菌素,是在多杀霉素的结构上将鼠李糖基3′位由甲氧基转变为乙氧基,且主链 C5~C6位存在两种异构体。多杀霉素由陶氏益农公司于1997年首次在美国登记,2007年该公司又推出乙基多杀菌素。目前,已有关于丁烯基多杀菌素的报道(尚未上市)。我国于1998年首次引进多杀霉素,随着该药专利到期,国内多家企业已成功合成并登记该药。
合成技术决定着多杀菌素类产品的种类。由于多杀霉素结构复杂,化学合成较难实现,所以使用刺糖多胞菌发酵产生多杀菌素类化合物是目前最重要的生产途径。在刺糖多胞菌中,涉及多杀菌素类化合物生物合成的基因共有23个,包括5个聚酮合成酶基因、4个聚酮桥接基因、8个糖生物合成基因、2个糖基转移酶基因和4个糖甲基化基因。通过对这些基因进行编辑可实现对多杀菌素类天然化合物化学结构的改造,但在目前合成的1,000种以上多杀菌素类似物中,未找到与天然多杀菌素A活性相当的类似物。我国在编辑聚酮合成酶基因改造多杀霉素方面取得了突破性进展,Song等开发了RedEx基因编辑方法,改造发酵菌株获得了高活性的丁烯基多杀菌素,为我国开发新型多杀菌素类杀虫剂奠定了基础。多杀菌素生物合成路径复杂,致使其难以异源表达,但随着基因编辑技术的发展,生物合成技术的成熟,未来会有更多的多杀菌素类杀虫剂出现。
1.2 作用方式
多杀菌素类杀虫剂属于大环内酯类杀虫剂,以触杀和胃毒作用为主。多杀霉素对害虫的卵、幼虫和成虫均有较高活性,且具有一定的选择性。该类杀虫剂属低毒农药,对天敌昆虫、哺乳动物、水生动物和鸟类安全。原药对雌性大鼠急性经口 LD50>5,000 mg/kg,对小鼠急性经口LD50>5,000 mg/kg,在环境中的半衰期3~5 d,无富集作用,不污染环境。研究发现,多杀霉素对红火蚁的触杀LC50 为3.44 mg/kg,胃毒 LD50 为4.22 mg/kg。徐志红等用点滴法、浸渍法和含药饲料喂食法证明了多杀菌素A对玉米螟具有触杀活性,对家蝇有触杀和胃毒活性。李丽荣发现多杀菌素类杀虫剂对花蓟马的胃毒活性高于触杀活性。
多杀菌素类杀虫剂主要作用于害虫的神经系统,通过刺激中枢神经系统导致其肌肉收缩、颤抖、衰竭,最后瘫痪死亡。注射多杀霉素后的蟑螂除表现上述症状外,还表现出由吞咽空气引起的腹部肿胀。类似的肌肉异常收缩症状在其他昆虫如烟草夜蛾幼虫、果蝇、家蝇上亦有所表现。烟草夜蛾幼虫暴露在多杀菌素类杀虫剂中,通常会仰面倒下、虫体持续震颤。田间观察发现,传统的杀虫剂通常导致害虫痉挛,较快便从叶片上掉落,而多杀霉素虽使害虫丧失活动能力,但害虫在作物叶片上停留时间较长。
02 多杀菌素类杀虫剂在国内应用现状
多杀霉素具有广泛的杀虫谱和高效的杀虫活性。截至2025年6月,中国农药信息网显示,国内登记的多杀霉素产品有152个,主要用于鳞翅目、缨翅目及双翅目害虫的防治;乙基多杀菌素登记产品较少, 共有5个(原药2个、制剂3个)。目前,除仓储害虫与卫生害虫外,两种杀虫剂登记的蔬菜作物共9种,防治对象包括蓟马、小菜蛾、甜菜夜蛾、斜纹夜蛾、豆荚螟、美洲斑潜蝇和蚜虫;登记的瓜果作物共4种,防治对象包括果蝇、橘小实蝇、橘大实蝇、蓟马和蚜虫;登记的大田作物共2种,防治对象为草地贪夜蛾、稻纵卷叶螟、二化螟和棉铃虫。
近年来,国内涉及多杀霉素和乙基多杀菌素的室内生测、田间防效报道较多。谷莉莉等开展施药防治设施白菜上小菜蛾的试验,调查得出,60 g/L乙基多杀菌素悬浮剂在施用后3 d和7 d的校正防效分别为96.91%和91.74%,显著高于对照药剂10%高效氯氰菊酯乳油的同期防效(69.35%和66.96%)。黄文枫等研究发现60 g/L乙基多杀菌素悬浮剂对白萝卜田黄曲条跳甲的防效较高,药后7 d防效达80%以上。除用于蔬菜害虫防治外,何超等发现乙基多杀菌素对石榴害虫井上蛀果斑螟的卵和幼虫有较强活性。
03 多杀菌素类杀虫剂的作用靶标
多杀菌素类杀虫剂的直接作用受体为烟碱型乙酰胆碱受体(nicotinic acetylcholine receptors,nAChRs)。Salgado等发现多杀霉素通过刺激昆虫神经系统,引起肌肉无意识收缩,导致昆虫过度兴奋,致使其疲劳直至衰竭。不同于一般的大环内酯类化合物,多杀菌素类杀虫剂具有独特的化学结构,决定了其独特的作用机制。作为一种神经毒性化合物,与竞争性结合抑制烟碱型乙酰胆碱受体的新烟碱类杀虫剂相比,多杀菌素类杀虫剂结合位点不同,是通过变构激动神经系统中的烟碱型乙酰胆碱受体α6亚基而发挥杀虫作用。
3.1 烟碱型乙酰胆碱受体的结构
烟碱型乙酰胆碱受体(nAChRs)属于半胱氨酸环配体门控离子通道超家族(Cys-loop LGIC superfamily),是烟碱类、新烟碱类、介离子类、多杀菌素类、亚砜亚胺类、丁烯酸内酯类、吡啶亚基类杀虫剂的主要作用靶标。目前,作用于该靶标的杀虫剂在全球化学杀虫剂市场中占比最高。昆虫nAChR结构复杂,是一类五聚体蛋白,包括α和β两种亚基。与哺乳动物相比,昆虫nAChR家族较小,虫体内通常存在1~11个α亚基、1~3个β亚基,但这些亚基基因通过可变剪切和RNA编辑增加了nAChR多样性。目前基于亚基的研究主要集中于受体功能、药剂开发、抗性机制等领域。昆虫基因组测序的进展显著促进了nAChR家族的鉴定和研究。目前,果蝇、家蚕、赤拟谷盗、冈比亚按蚊等多种昆虫的nAChR基因家族已被鉴定与分析,这为新型杀虫剂的靶点设计和抗药性管理提供了理论依据。
20世纪80年代,石纹电鳐nAChR亚基的成功克隆,为研究脊椎动物nAChR结构奠定了基础。基于该模型的亲和标记和结构分析表明,脊椎动物nAChR是由5个亚基组成的290kDa跨膜蛋白复合体,其激动剂结合位点位于相邻亚基的结合面处,并具有阳离子选择性。随后的研究发现,所有nAChR亚基在结构上具有一致性,包括胞外N端约200个氨基酸保守区域,4个保守跨膜区域和跨膜区3和4之间的氨基酸可变胞内大环区域(图1)。各亚基在N端胞外区成环形排列,包括LoopA~F共6环;在Loop E和Loop B之间,有1个被13个氨基酸间隔的半胱氨酸环(Cys-Loop),在氨基酸装配和离子通道门控的动力学中发挥重要作用。根据亚基组成特征,nAChR可分为α同源同型五聚体、α异源同型五聚体、α异源异型五聚体和非α异源异型五聚体。不同亚型受体对乙酰胆碱和阳离子的亲和力存在差异,且其在昆虫不同发育阶段和不同组织中的表达也存在差异,可能与昆虫在不同时期对环境的感知力及对杀虫剂的敏感性存在相关性。
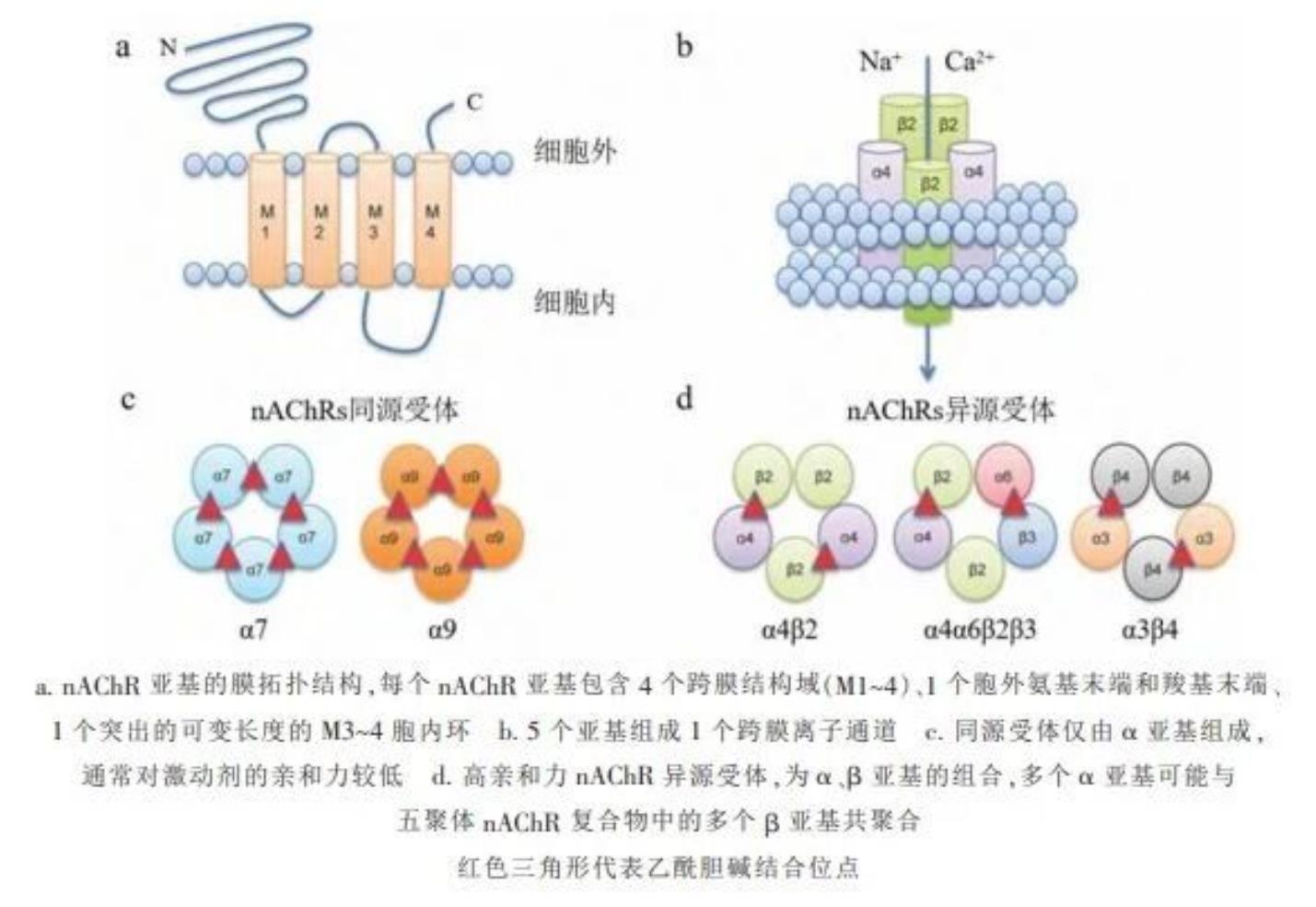
图1 烟碱型乙酰胆碱受体(nAChRs)结构图
3.2 烟碱型乙酰胆碱受体的分类与命名
烟碱型乙酰胆碱受体亚基分类的主要依据在于亚基N端结构域Loop C上是否有两个相连的半胱氨酸Cys残基,若有则定名为α亚基,若无则定名为非α亚基。根据目前已鉴定的nAChR基因,昆虫烟碱型乙酰胆碱受体包含α和β两种亚型。
构成乙酰胆碱受体的亚型由组成它的亚基数量命名,例如(α4)2(β2)3 表示该亚型由2个α4和 3个β2组成。当研究中不清楚亚型的具体组成时,可能会使用星号加以标注,例如α4β1*表示该亚型组成有α4和β1,但同时可能还包含其他亚基类型。由此,烟碱型乙酰胆碱受体分为由同一亚基组成的同型五聚体和由不同亚基组成的异型五聚体。异型五聚体又因组成亚基是否同源分为同源异型五聚体(由不同的α亚基组成)和非同源异型五聚体(由α和非α亚基组成)。
3.3 烟碱型乙酰胆碱受体的功能
烟碱型乙酰胆碱受体必须通过多亚基形成聚合体的方式才能在生物体内行使相应的功能,其广泛分布于中枢和外周神经系统中,参与调控神经递质的释放、细胞的兴奋传导等生理过程。对于昆虫nAChRs的研究,主要关注其在神经信号传递和抗杀虫剂靶点研究方面。昆虫nAChRs与脊椎动物的nAChRs在结构和功能上存在差异,这使得它们成为开发特异性杀虫剂的潜在目标。植物生物碱烟碱和真菌抗生素多杀霉素均对昆虫的nAChRs具有高亲和力,能有效干扰昆虫的神经系统,从而发挥显著的杀虫效果。在害虫防治领域,还需进一步探索nAChRs的亚基组成、配体结合特性及其在不同生物体中的特异性功能,以便更好地利用这些受体发挥综合防控的作用。目前对于该受体的研究方法涵盖了电生理学、分子生物学、结构生物学及行为学方法,旨在深入理解受体的功能和作用机制。尽管取得了一定的进展,但仍面临着亚型组成复杂、体外表达困难以及受体结构和功能动态变化难以捕捉等问题。此外,昆虫nAChRs对于不同种类的农药表现出敏感性差异,反映了其在进化上的多样性和复杂性,这为相关研究提出了挑战,同时也带来了新的方向。未来需要更深入地探索昆虫nAChRs结构与功能的关系、亚型特异性的信号传导路径以及它们如何影响昆虫对环境的适应性和生存策略,这对于开发更为安全、有效的害虫防治策略具有重要意义。
nAChRs的表达模式及其生物学功能的研究已覆盖多个昆虫种类,包括重要的经济害虫和模式生物。 刘少凯等对棉铃虫的研究表明,nAChR α7在昆虫不同发育阶段和不同组织中表现出特定的表达模式,这些表达变化与昆虫的生长发育、神经信号传导和抗药性形成密切相关;Xu等研究发现,nAChRs在二化螟不同组织中的表达存在一定差异,可能与其亚基功能相关。对黑腹果蝇的研究表明,nAChR亚基在昆虫不同发育阶段承担着与逃逸、睡眠、指导神经突出发育相关的多项功能。有研究使用CRISPR/Cas9技术敲除了棉铃虫和小菜蛾的nAChRα6亚基基因后,可导致其对多杀霉素产生高水平抗性。有研究者进一步发现, nAChR α6亚基基因的突变是试虫产生抗药性的主要原因。Lu等研究发现果蝇nAChR不同亚基基因突变后,其对杀虫剂表现出不同抗性,以α6亚基突变体对多杀霉素的抗性最显著,其他亚基突变体对多杀霉素也表现出一定抗性。
04 讨 论
多杀菌素类杀虫剂在落叶果树害虫防治中的应用较少,Perry等认为,丁烯基多杀菌素对苹果蠹蛾的防效显著高于多杀霉素和乙基多杀菌素,笔者所在团队近期研究发现,多杀菌素对落叶果树重大害虫桃小食心虫的卵具有较高活性。该类杀虫剂是否适用于果树食心虫的防治,需要开展田间防治试验评价其应用效果;对于果树其他害虫是否有效 也需进一步探索。
昆虫nAChRs是多种杀虫剂的靶标,药理学特性较为复杂。有报道提出,多杀菌素类杀虫剂除nAChRs外可能还存在另一作用靶标——GABA门控氯离子通道,并推测该类药剂对两种位点的调控是其具有较强杀虫活性的原因。多杀霉素对桃小食心虫的高活性是否与上述两个位点的调控有关,还需进一步研究。随着分子生物学、合成生物学、计算化学等学科技术的交叉融合,对昆虫nAChRs结构及功能机制的解析将不断深入,这将为创制新型高效、环境友好的绿色杀虫剂提供更广阔的思路。随着我国在农药创新领域科技能力的持续提升,相信未来将开发出具有自主知识产权的新一代多杀菌素类杀虫剂品种。